
In vitro Anti-Inflammatory Potency of Sanguinarine and Chelerythrine via Interaction with Glucocorticoid Receptor
- DOI
- 10.2991/efood.k.210118.001How to use a DOI?
- Keywords
- Anti-inflammatory potency; sanguinarine; chelerythrine; glucocorticoid receptor
- Abstract
As quaternary benzo[c]phenanthridine alkaloids, sanguinarine and chelerythrine possess multiple biological activities including anti-inflammatory effects. They have been confirmed to regulate gene expression of inflammatory-related cytokines at transcription level. This work aims to investigate the glucocorticoid-like anti-inflammatory effects of sanguinarine and chelerythrine, which may be mediated by Glucocorticoid Receptor (GR). Firstly, the interactions of GR with sanguinarine and chelerythrine were investigated by fluorescence polarization assay and the result showed that these two alkaloids could bind to GR with potent affinities. Luciferase reporter assay showed that sanguinarine was a weak GR agonist but chelerythrine failed to induce GR transcriptional activation in HeLa cells. These two alkaloids at high concentration showed similar tumor necrosis factor-α inhibitory potency to dexamethasone in RAW 264.7 cells, indicating their anti-inflammatory effects in vitro. In addition, the results of molecular docking and dynamics simulations showed that sanguinarine and chelerythrine could bind effectively with GR and might serve as its functional ligands. In conclusion, chelerythrine was speculated to be a selective GR modulator that might exhibit GR-mediated beneficial actions with reduced side effects.
- Graphical Abstract
- Copyright
- © 2021 The Authors. Publishing services by Atlantis Press International B.V.
- Open Access
- This is an open access article distributed under the CC BY-NC 4.0 license (http://creativecommons.org/licenses/by-nc/4.0/).
1. INTRODUCTION
As quaternary benzo[c]phenanthridine alkaloids, sanguinarine and chelerythrine can be isolated from Sanguinaria canadensis, Chelidonium majus, and Macleaya cordata [1,2]. These two alkaloids possess multiple biological activities including anti-inflammatory, anti-microbial, and anti-tumor effects [3]. Furthermore, they have also been reported to inhibit human liver cytochrome P450 enzymes and rat liver aromatic amino acid decarboxylase [4–6]. Over the past few decades, sanguinarine and chelerythrine have been widely used for the prevention and treatment of various diseases due to their low toxicity and high biocompatibility [7]. A series of studies suggest that they might be promising drugs to enhance sensitivity and overcome therapeutic resistance in both human ovarian cancer cells and xenograft tumors [8,9]. Although sanguinarine and chelerythrine have been confirmed to regulate gene expression of inflammatory-related cytokines at transcription level [10,11], the underlying mechanisms of their anti-inflammatory effects are still largely unclear.
In clinical practice, the synthetic glucocorticoids such as Dexamethasone (DEX) have been widely used as anti-inflammatory agents. The in vitro and in vivo anti-inflammatory effects of DEX have been confirmed since 1948 [12,13]. DEX exerts a regulatory role in inflammation via binding to Glucocorticoid Receptor (GR), which is a ligand-activated transcription factor that belongs to nuclear receptor superfamily [14,15]. After binding with ligands such as DEX, GR can translocate from the cytoplasm to the nucleus and then play a relevant role in inflammation and immunity [16]. Unfortunately, a series of glucocorticoid-associated side effects including osteoporosis, diabetes, depression, and muscle wasting have been observed in clinical practice [17]. It has been reported that the side effects induced by glucocorticoids are attributed to transactivation of GR responsive genes [18,19]. Meanwhile, the clinical effects of glucocorticoids are primarily mediated via transrepression of expression of relevant pro-inflammatory genes [20]. Therefore, there is an urgent need for screening of Selective Glucocorticoid Receptor Modulators (SEGRMs) that can separate transactivation from transrepression of GR responsive genes.
Numerous phytochemicals derived from fruits, vegetables, and food legumes have been confirmed for their anti-inflammatory effects [21]. Among which, Compound A (CpdA) is a natural phytochemical that has been identified as a SEGRM [22]. Possessing dissociated characteristics, CpdA obtains a better balance between transactivation and transrepression of GR responsive genes, resulting in its advantage as an anti-inflammatory agent with reduced side effects. In addition to CpdA, several other phytochemicals also reduce inflammation with reduced side effects, serving as SEGRMs [23]. Based on the chemical structures of sanguinarine and chelerythrine (Figure 1), these two alkaloids were postulated to exhibit glucocorticoid-like anti-inflammatory effects via binding to GR. In this work, the in vitro anti-inflammatory potency of sanguinarine and chelerythrine was assessed and their interactions with GR were also investigated.

Structures of sanguinarine (left) and chelerythrine (right).
2. MATERIALS AND METHODS
2.1. Materials
All the cell lines were purchased from the Cell Bank of the Chinese Academy of Sciences (Shanghai, China). The Glucocorticoid Receptor Ligand Binding Domain (GR-LBD) recombinant human protein, DEX fluorescein, and Lipofectamine 2000 transfection reagent were purchased from Thermo Fisher Scientific (San Jose, CA, USA). Dimethylsulfoxide (DMSO), Lipopolysaccharide (LPS), and DEX were purchased from Sigma-Aldrich (St. Louis, MO, USA). Sanguinarine and chelerythrine were purchased from Yuanye Biotechnology Co., Ltd. (Shanghai, China). Fetal Bovine Serum (FBS), penicillin–streptomycin and trypsin–ethylenediaminetetraacetic acid solution were purchased from Gibco (Grand Island, NY, USA). The Enzyme-linked Immunosorbent Assay (ELISA) kit for mouse Tumor Necrosis Factor-α (TNF-α) was purchased from Yunkelong Technology Co., Ltd. (Wuhan, China). Phosphate-buffered Saline (PBS) and Dulbecco’s Modified Eagle’s Medium (DMEM) were purchased from Biological Industries (Beit Haemek, Israel). All other reagents were of analytical grade.
2.2. Fluorescence Polarization Competitive Binding Assay
Fluorescence polarization competitive binding assay was performed by using a method that was previously described [24]. DEX fluorescein was used as a tracer, which can be displaced from GR-LBD in presence of competitive ligands. The final DMSO concentration was kept below 1% (v/v) in each experiment. The plate was incubated at 4°C and then the fluorescence polarization value of each well was measured on a microplate reader Tecan Infinite M1000Pro (Tecan Austria GmbH, Grödig, Austria). The excitation and emission wavelength were set at 485 and 535 nm, respectively. Each alkaloid was measured in triplicate. Statistical analyses and curve fitting were performed using GraphPad Prism (version 5, GraphPad Software, La Jolla, CA, USA). Data were presented as mean ± SEM.
2.3. Luciferase Reporter Assay
HeLa cells with a density of 105 cells per well were seeded for 18 h and then were transiently transfected with plasmids, including expression plasmid pcDNA3.1(+)-hGR, GR-driven reporter plasmid GRE-Luc, and internal control plasmid pRL-TK. The transfection assay was performed with Lipofectamine 2000 transfection reagent [25]. After 24 h of transfection, HeLa cells were treated with DMSO, the GR agonist DEX (100 nM), or the different concentrations of alkaloid for another 24 h. The firefly luciferase activity was determined and normalized against to that of Renilla by a Dual-Glo Luciferase Assay System (Promega, Madison, WI, USA). Each alkaloid was measured in triplicate on a microplate reader SpectraMax i3x (Molecular Devices, Sunnyvale, CA, USA), and the GR agonist DEX was used as a positive control.
2.4. Cytokine Assay
Lipopolysaccharide-induced inflammation assay was performed in RAW 264.7 cells. Briefly, RAW 264.7 cells were seeded at a density of 5 × 105 in 12-well plates and cultured in DMEM supplemented with 10% FBS at 37°C with 5% CO2. The cells in 12-well plates were washed twice with sterile PBS and then the fresh medium was added. Subsequently, LPS (1 μg/mL) was added to the fresh medium and incubated for 2 h. Sanguinarine and chelerythrine were added to the culture medium in final concentrations of 0.1, 0.5, 1, 2, 4, 5 μM and 0.1, 0.2, 0.5, 1, 2, 5 μM, respectively. After incubated for 12 h, the culture medium was harvested and centrifuged to clear cellular debris. Cell supernatants were assayed for pro-inflammatory cytokine TNF-α production by ELISA [26]. DEX (10 μM) was used as a positive control.
2.5. Molecular Docking
The protein data of GR-LBD was provided by Protein Data Bank with ID 4UDC [27]. Chimera was used to preprocess the protein and to remove ligand DEX as well as water molecules from the complex [28]. AutoDockTools was used to add the hydrogen atoms. The structures of sanguinarine and chelerythrine were generated by GaussView and their energy minimization was performed by Gaussian 09W. AutoDockTools was used to dock sanguinarine and chelerythrine into GR-LBD with 10 independent runs. The predicted binding poses of sanguinarine and chelerythrine with lowest binding energies were used for sequential analysis.
2.6. Molecular Dynamics Simulations
GROMACS was used to assess the binding stabilities of sanguinarine and chelerythrine with GR-LBD. Firstly, CHARMM36 all-atom force field was used to prepare the topology of GR-LBD [29]. The topologies of sanguinarine and chelerythrine were obtained by CGenFF server. The alkaloid-GR-LBD complex was placed in a cubic box consisting simple point charge water. Sodium ions were added to maintain the electrostatic neutrality. After energy minimization, the system was equilibrated within constant number of particles-volume-temperature (NVT) and constant number of particles-pressure-temperature (NPT) ensembles. The Root Mean Squared Deviation (RMSD) values for alkaloid-GR-LBD complex were calculated to assess their binding stabilities during 20 ns dynamics simulation.
3. RESULTS AND DISCUSSION
3.1. GR Binding Affinities of Sanguinarine and Chelerythrine
In this work, DEX fluorescein was used as a tracer for fluorescence polarization assay. At the beginning of fluorescence polarization assay, DEX fluorescein and GR-LBD recombinant protein form a complex that rotates slowly, producing a high polarization value. DEX fluorescein can be displaced from GR-LBD in presence of competitive ligands, producing a low polarization value. Hence, the interactions between GR-LBD and its ligands could be monitored via determining the change of fluorescence polarization signal. The competitive binding curves of these two alkaloids were shown in Figure 2 and their corresponding IC50 values were listed in Table 1. Sanguinarine and chelerythrine exhibit concentration-dependent binding to GR-LBD, with IC50 values of 4.30 × 10−9 and 8.01 × 10−13 mg/mL, respectively. The result of fluorescence polarization assay showed that both sanguinarine and chelerythrine could bind to GR with potent affinities. Therefore, it can be speculated that sanguinarine and chelerythrine might be functional ligands for GR.
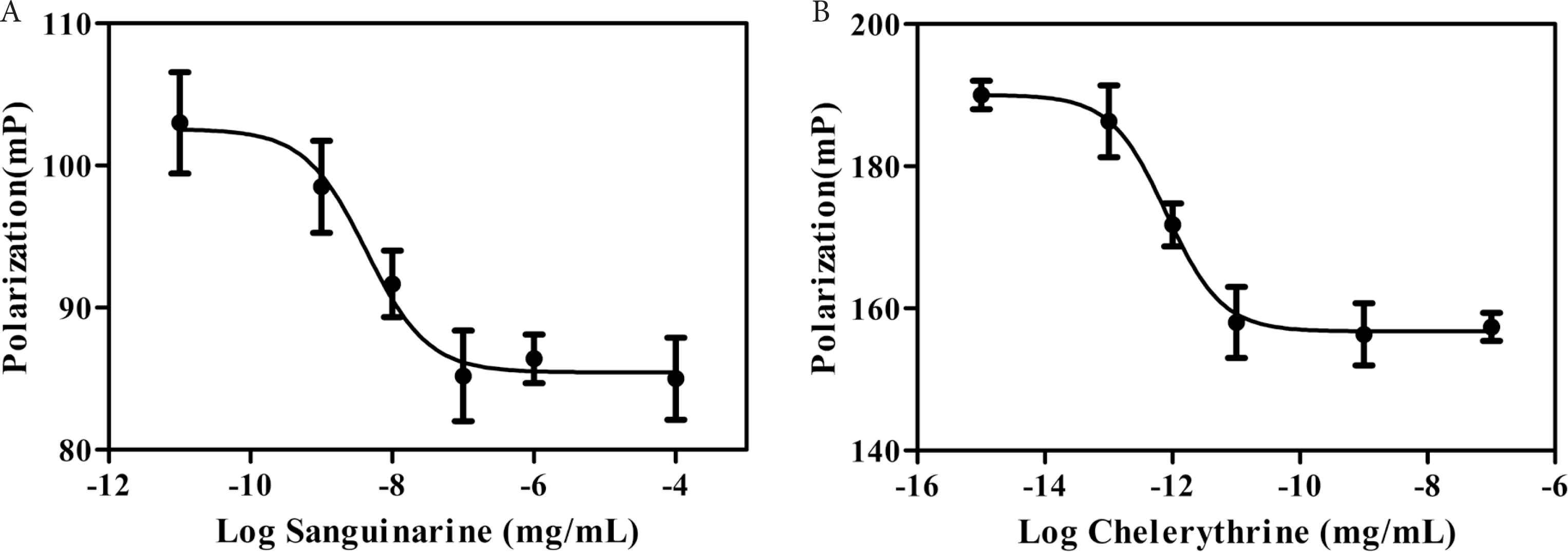
Fluorescence polarization competitive binding assay for sanguinarine (A) and chelerythrine (B).
| Compound | CSEV (Å3) | IC50 (mg/mL) | Hydrogen bonds | Hydrophobic contacts | Binding energy (kcal/mol) |
|---|---|---|---|---|---|
| Sanguinarine | 249.59 | 4.30 × 10−9 | Asn564 and Thr739 | Met560, Leu563, Met604, Ala605, Leu608, Met646, Leu732, and Ile747 | −8.75 |
| Chelerythrine | 279.84 | 8.01 × 10−13 | Gln570 and Arg611 | Leu563, Leu566, Met601, Met604, Ala605, Leu608, Phe623, Met646, Leu732, and Phe749 | −8.87 |
The Connolly Solvent-Excluded Volume (CSEV) of sanguinarine and chelerythrine, as well as their IC50 values in the fluorescence polarization assay and docking results with GR-LBD
3.2. GR Transcriptional Activities of Sanguinarine and Chelerythrine
As shown in Figure 3, the agonist DEX significantly induces transcriptional activation of GR at the concentration of 100 nM, indicating that this reporter gene system is suitable for assessing the GR transcriptional activities of sanguinarine and chelerythrine. It can be observed that sanguinarine also slightly induces transcriptional activation of GR at the high tested concentrations (Figure 3), indicating that sanguinarine is a weak GR agonist, although not as potent as DEX. On the contrary, chelerythrine does not induce transcriptional activation of GR at the tested concentrations ranging from 0.1 nM to 1 μM (Figure 3), indicating that chelerythrine show no agonistic activities toward GR. Since the aforementioned fluorescence polarization assay has confirmed the GR binding affinity of chelerythrine, we can conclude that chelerythrine is able to bind to GR but fail to induce GR transcriptional activation. It has been reported that the side effects caused by glucocorticoids are mainly due to the transactivation of GR responsive genes [30]. Several synthetic and natural compounds have been characterized as dissociated agonists for GR, with the advantages of altering the balance between transactivation and transrepression of GR [23]. Hence, it can be speculated that chelerythrine might exhibit GR-mediated beneficial actions with reduced side effects.
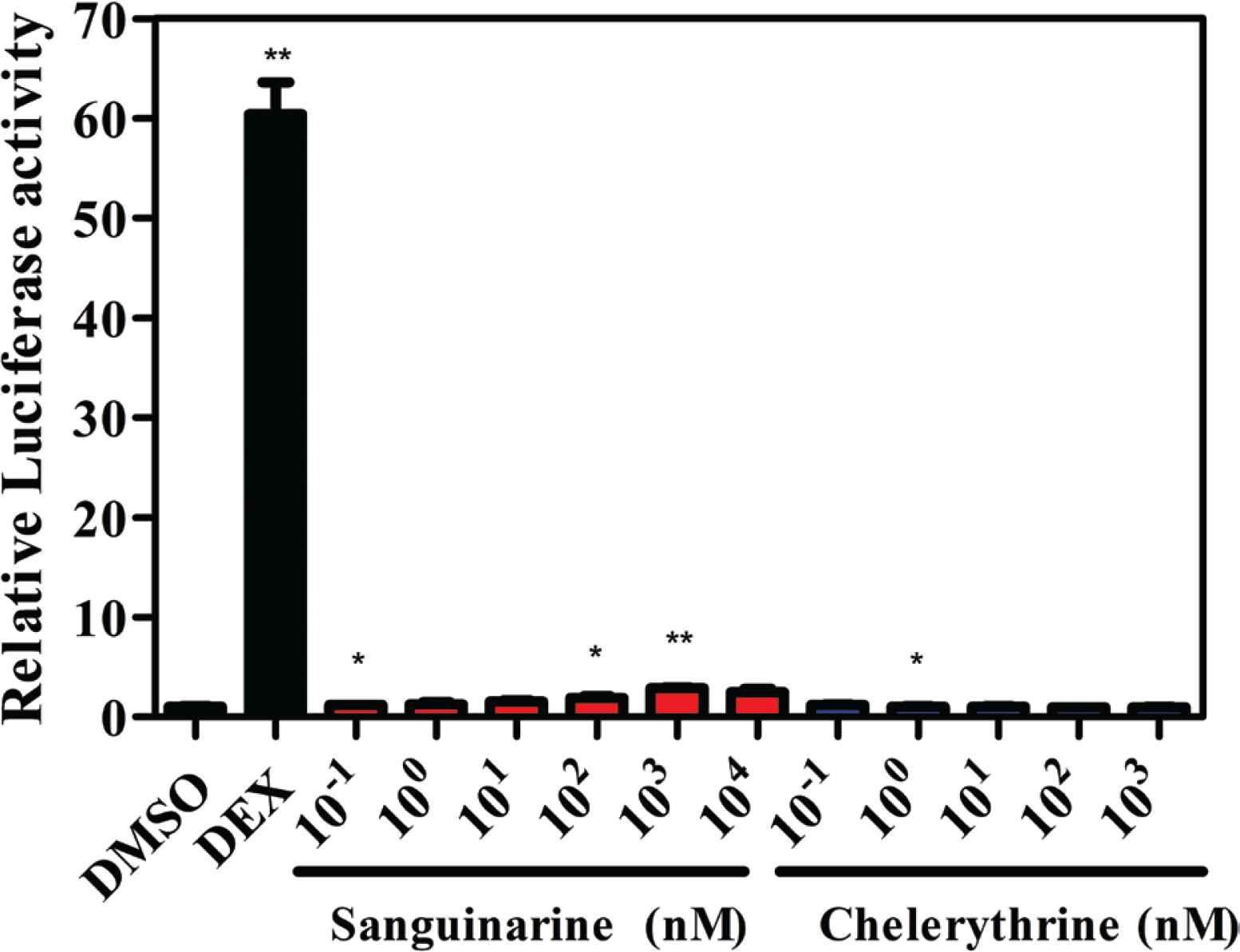
Luciferase reporter assay for sanguinarine and chelerythrine in HeLa cells. DEX, dexamethasone (100 nM). *p < 0.05 and **p < 0.01 compared with the control group (DMSO).
3.3. Effects of Sanguinarine and Chelerythrine on the Production of TNF-α
In order to investigate the in vitro anti-inflammatory potency of sanguinarine and chelerythrine, this work focuses on the regulation of protein expression of inflammatory-related cytokine by these two alkaloids. After stimulated with LPS for 2 h, the production of pro-inflammatory cytokine TNF-α increases significantly in RAW 264.7 cells (Figure 4). DEX at the concentration of 10 μM inhibits LPS-induced TNF-α production. It can be observed that sanguinarine and chelerythrine also inhibit LPS-induced TNF-α production and exhibit a concentration-dependent manner. Herein, these two alkaloids at concentration of 5 μM show similar inhibitory potency to DEX, indicating their anti-inflammatory effects in vitro.
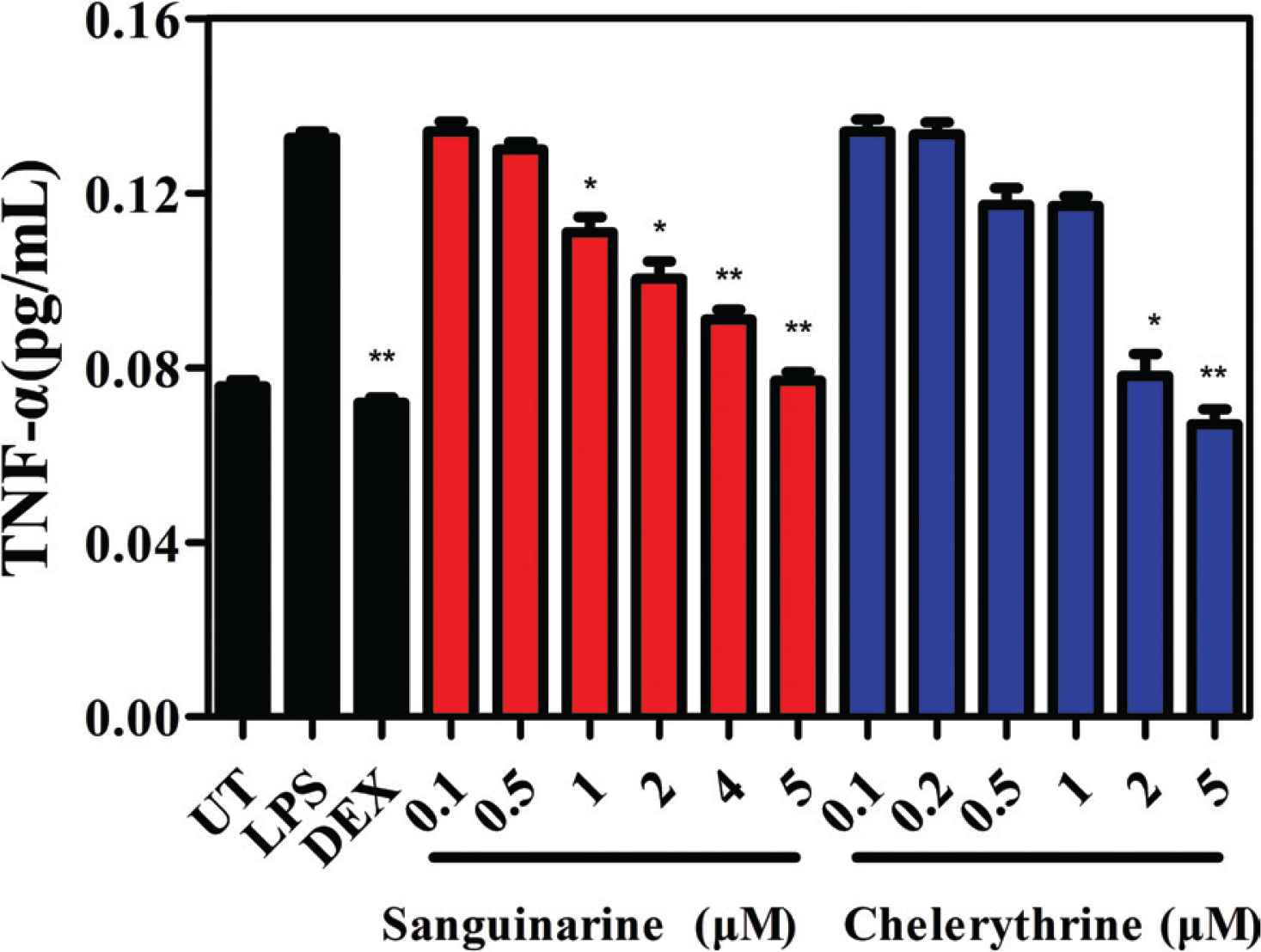
Effects of sanguinarine and chelerythrine on the production of TNF-α in RAW 264.7 cells. UT, untreated; LPS, lipopolysaccharide; DEX, dexamethasone (10 μM). *p < 0.05 and **p < 0.01 compared with the LPS-treated group.
3.4. Binding Poses of Sanguinarine and Chelerythrine with GR-LBD
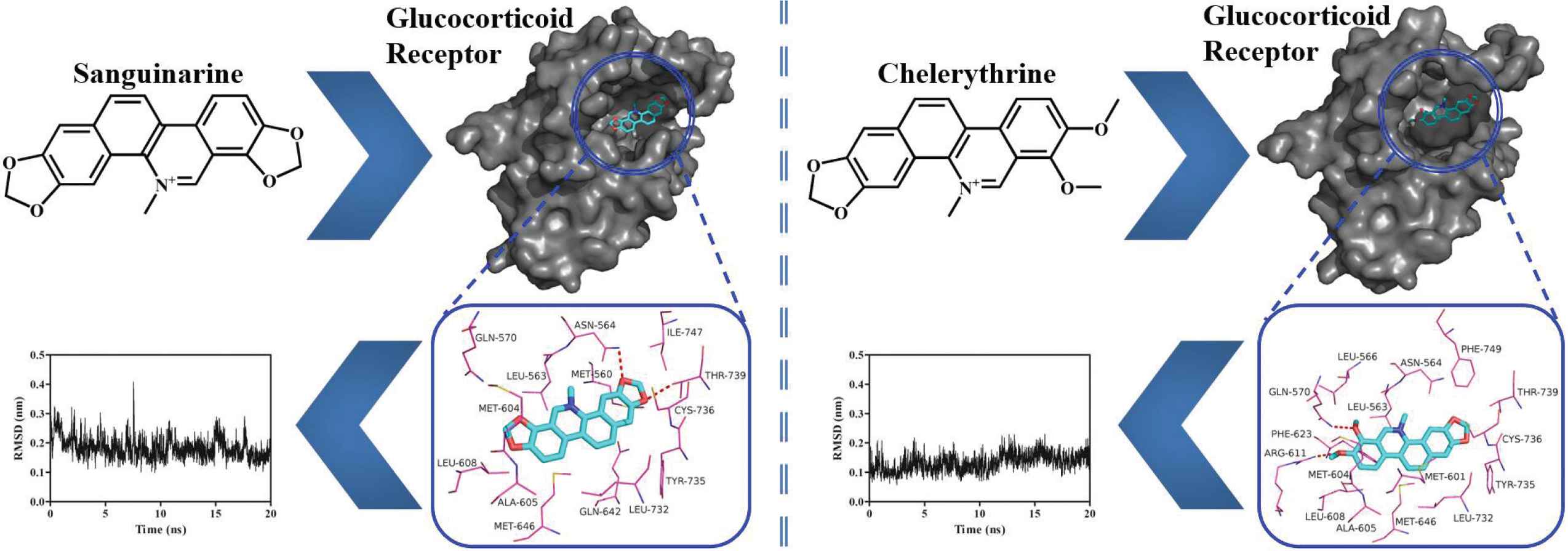
As shown in Figure 5A and 5B, sanguinarine and chelerythrine can fit into the hydrophobic cavity of GR. As shown in Figure 5C and 5D, these two alkaloids are surrounded by both hydrophobic and hydrophilic residues. Sanguinarine is stabilized via the hydrogen-bonding interactions with residues Asn564 (2.64 Å) and Thr739 (2.72 Å), while chelerythrine is stabilized via the hydrogen-bonding interactions with residues Gln570 (2.75 Å) and Arg611 (2.81 Å), as listed in Table 1. Met560, Leu563, Met604, Ala605, Leu608, Met646, Leu732, and Ile747 make hydrophobic interactions with sanguinarine, while Leu563, Leu566, Met601, Met604, Ala605, Leu608, Phe623, Met646, Leu732, and Phe749 make hydrophobic interactions with chelerythrine. As shown in Figure 1, sanguinarine contains two methylenedioxy rings, while chelerythrine only contains one methylenedioxy ring and two methoxy groups [31]. These slight structural differences of sanguinarine and chelerythrine result in their different binding affinities. As listed in Table 1, the Connolly solvent-excluded volume value of chelerythrine is larger than that of sanguinarine, resulting in more hydrophobic contacts of chelerythrine toward GR compared with sanguinarine, which leads to stronger binding energy of chelerythrine than sanguinarine. Interestingly, the order of the calculated binding energies in computer simulation is consistent with the experimentally determined binding affinities (IC50 values) in fluorescence polarization assay.
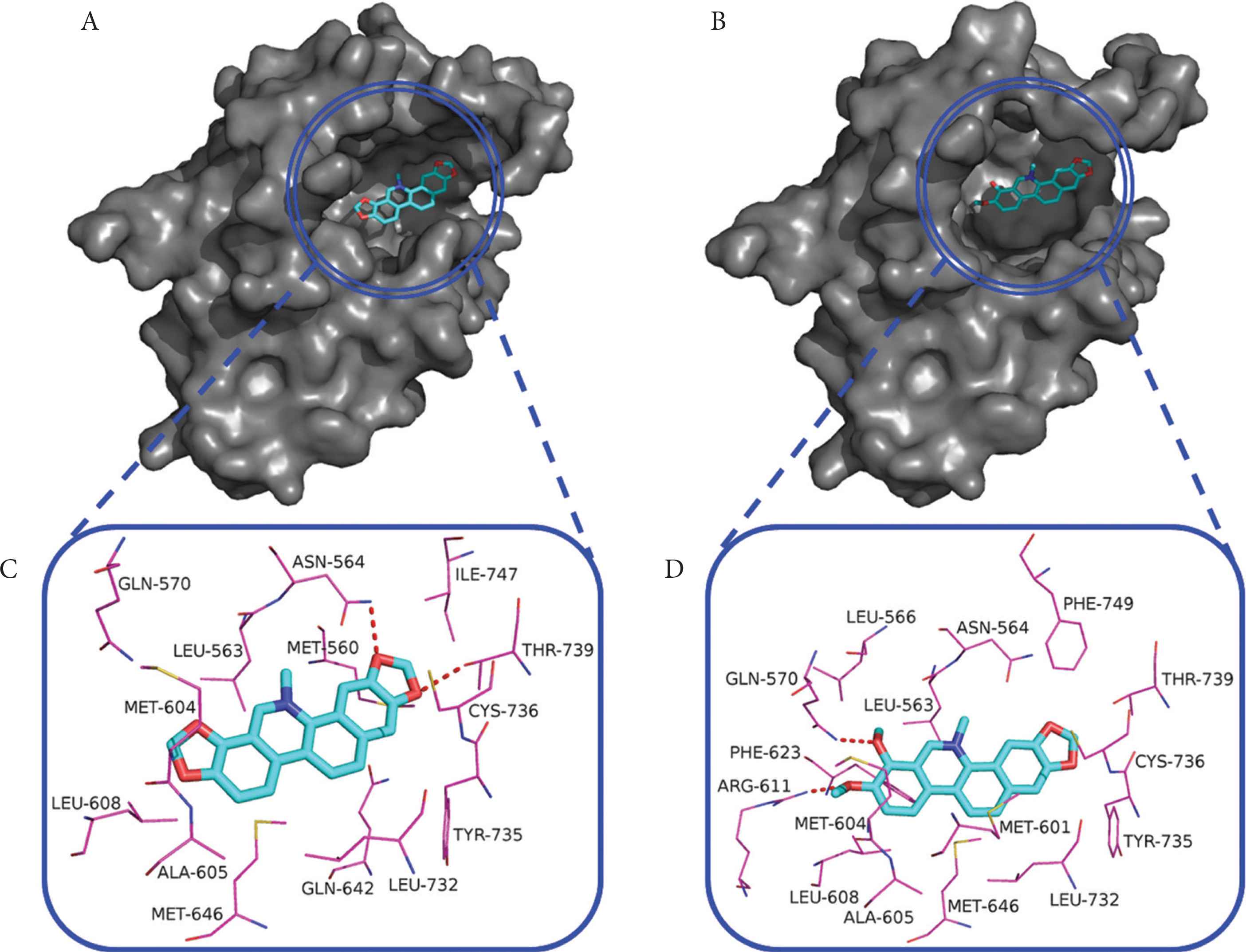
The binding poses of sanguinarine (A) and chelerythrine (B) in the hydrophobic cavity of GR-LBD and the interaction networks of sanguinarine (C) and chelerythrine (D) with residues lying within 4 Å distance. Red dashed lines, hydrogen bonds.
3.5. Binding Stabilities of Sanguinarine and Chelerythrine with GR-LBD
The binding stabilities of sanguinarine and chelerythrine with GR-LBD were assessed by molecular dynamics simulations and the calculated RMSD values can be seen in Figure 6. During 20 ns dynamics simulation, sanguinarine and chelerythrine basically achieve stability with the average RMSD values of 0.18 ± 0.04 nm (Figure 6A) and 0.13 ± 0.03 nm (Figure 6B), respectively. It is noteworthy that chelerythrine is more stable than sanguinarine, which may be due to the stronger binding energy of chelerythrine than sanguinarine (Table 1). Interestingly, both sanguinarine and chelerythrine induce a slight disturbance toward GR-LBD, with the average RMSD values of 0.11 ± 0.02 nm (Figure 6C) and 0.14 ± 0.01 nm (Figure 6D), respectively. In conclusion, sanguinarine and chelerythrine could bind effectively with GR to induce its nuclear translocation.
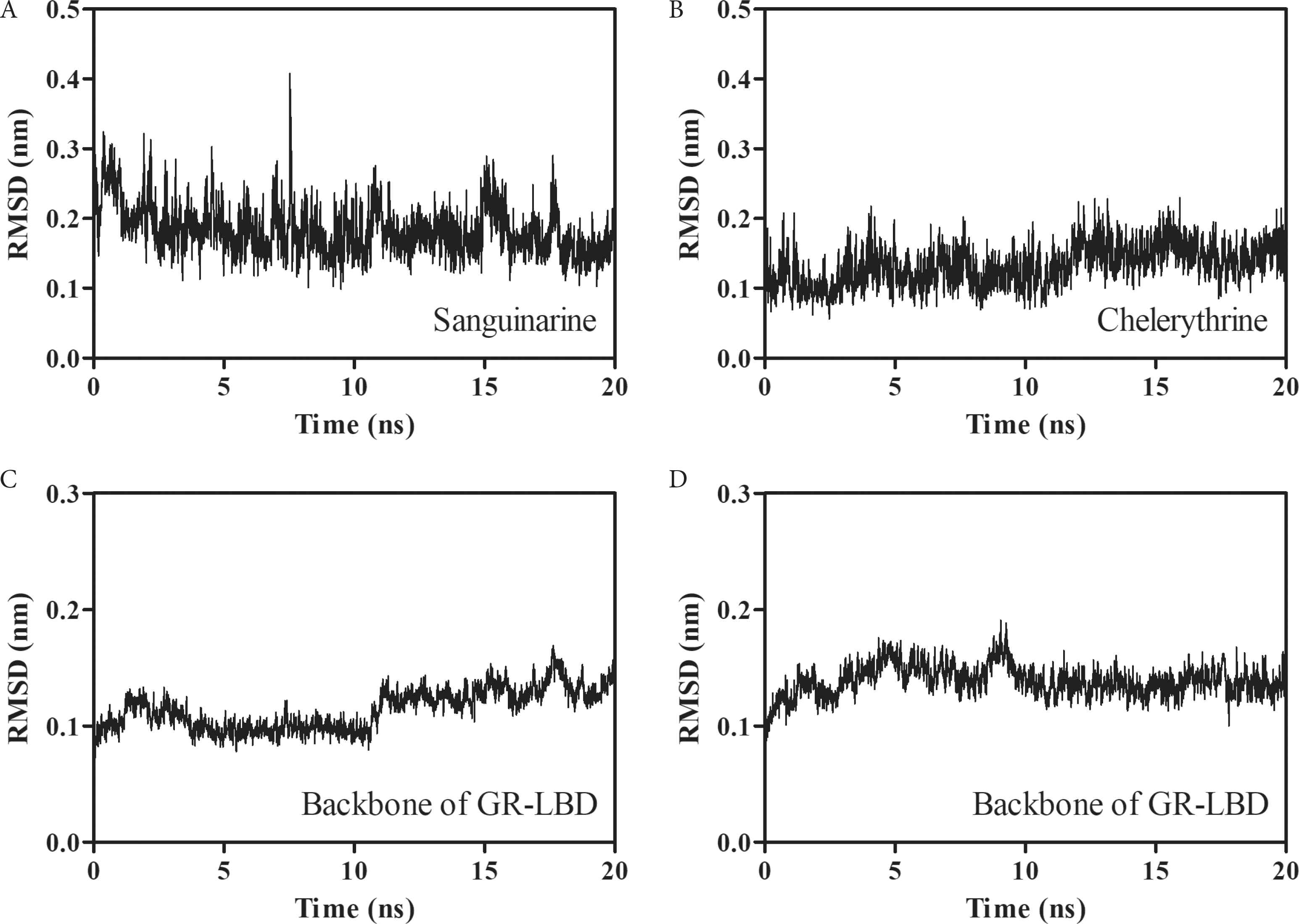
The variations of RMSD values for sanguinarine (A) and chelerythrine (B) as well as their respective backbones of GR-LBD (C and D).
4. CONCLUSION
In this work, the glucocorticoid-like anti-inflammatory effects of sanguinarine and chelerythrine was investigated. These two alkaloids could bind to GR with potent affinities. Interestingly, sanguinarine was a weak GR agonist but chelerythrine failed to induce GR transcriptional activation in HeLa cells. Both sanguinarine and chelerythrine at high concentration showed similar TNF-α inhibitory potency to DEX in RAW 264.7 cells, indicating their anti-inflammatory effects in vitro. The results of molecular docking and dynamics simulations showed that sanguinarine and chelerythrine could bind effectively with GR and might serve as its functional ligands. In conclusion, chelerythrine was speculated to be a selective GR modulator that might exhibit GR-mediated beneficial actions with reduced side effects.
CONFLICTS OF INTEREST
The authors declare they have no conflicts of interest.
AUTHORS’ CONTRIBUTION
JZ contributed in literature search and manuscript writing. YL contributed in experimental data collection and analyses. LR contributed in conceptualization and project administration. TZ contributed in supervision and manuscript review.
ACKNOWLEDGMENTS
This work was supported by
Footnotes
REFERENCES
Cite this article
TY - JOUR AU - Jie Zhang AU - Yuan Liang AU - Li Ren AU - Tiehua Zhang PY - 2021 DA - 2021/01/21 TI - In vitro Anti-Inflammatory Potency of Sanguinarine and Chelerythrine via Interaction with Glucocorticoid Receptor JO - eFood SP - 392 EP - 398 VL - 1 IS - 6 SN - 2666-3066 UR - https://doi.org/10.2991/efood.k.210118.001 DO - 10.2991/efood.k.210118.001 ID - Zhang2021 ER -