
Dendrobine Suppresses Lipopolysaccharide-induced Gut Inflammation in a Co-culture of Intestinal Epithelial Caco-2 Cells and RAW264.7 Macrophages
- DOI
- 10.2991/efood.k.210409.001How to use a DOI?
- Keywords
- Dendrobine; inflammatory bowel diseases; tight junctions; NF-κB pathway
- Abstract
We evaluated dendrobine for its potential effect on Inflammatory Bowel Disease (IBD) progression using the intestinal epithelial Caco-2/RAW264.7 macrophage co-culture cell model. The results showed that dendrobine maintained the tight junction (TJ) proteins in co-cultured Caco-2 cells, suggesting its protective effect on intestinal integrity. Moreover, the findings indicated that the dendrobine group downregulated levels of tumor necrosis factor-α (TNF-α), proinflammatory Interleukin (IL)-1β and IL-6, and inflammatory markers such as cyclooxygenase-2 (COX-2) and inducible nitric oxide synthase (iNOS) in co-cultured RAW264.7 cells. Additionally, dendrobine-mediated anti-inflammatory effects were observed based on the inhibition of LPS-induced proinflammatory mediators of nuclear factor kappa B (NF-κB)-related signaling. These results demonstrated the protective effect of dendrobine at effectively ameliorating intestinal barrier disruption and inhibiting activation of the NF-κB pathway in macrophages through the intestinal barrier.
- Graphical Abstract
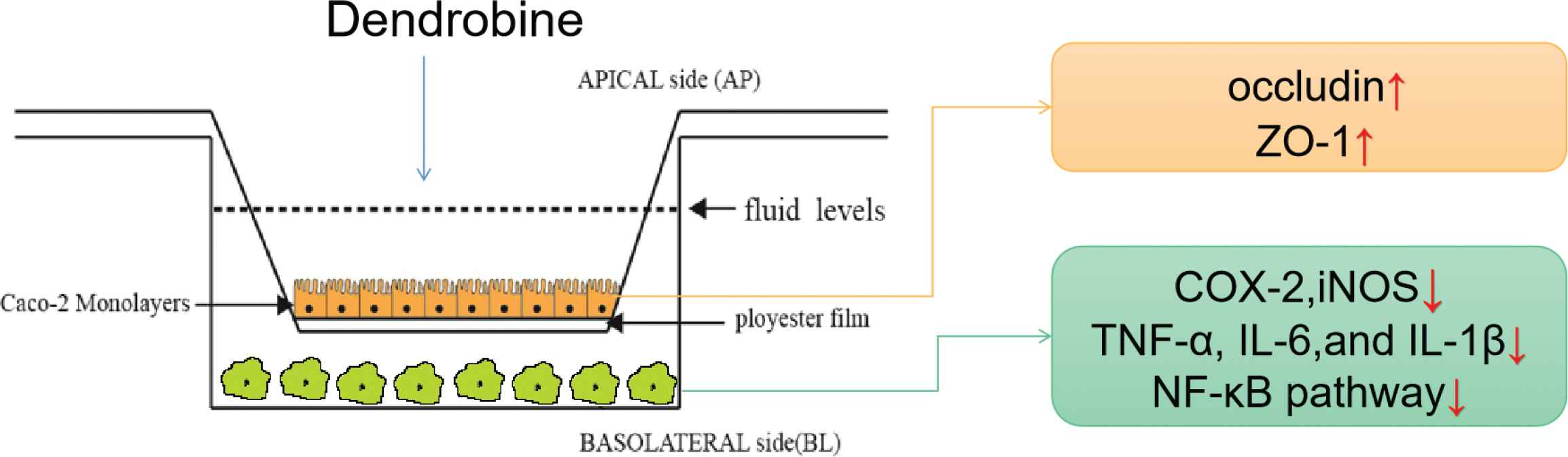
- Copyright
- © 2021 The Authors. Publishing services by Atlantis Press International B.V.
- Open Access
- This is an open access article distributed under the CC BY-NC 4.0 license (http://creativecommons.org/licenses/by-nc/4.0/).
1. INTRODUCTION
Inflammatory Bowel Disease (IBD) is a type of chronic inflammation marked by the abnormal activation of macrophages in the intestinal lamina propria [1]. Intestinal Epithelial Cells (IECs) are part of this barrier and prevent the diffusion of pathogens into the circulatory system [2]. If the intestinal barrier is disrupted, intestinal permeability increases and immune homeostasis is disturbed, leading to intestinal diseases [3]. The Tight Junction (TJ) proteins occludin and Zonula Occludens Protein 1 (ZO-1) participate in maintaining intestinal structural integrity [4]. The mechanism underlying IBD causes increased intestinal inflammation and TJ permeability, which is associated with increased levels of anti-inflammatory cytokines in the intestine and blood [5].
In vitro study of the intestinal epithelial barrier function requires the establishment and characterization of models reflecting the physiological relationships of cells. Many scholars have attempted to use co-culture models of IECs and immune cells to replicate their interactions in vitro [6]. The intestinal epithelial Caco-2 cell line has been widely used to simulate the intestinal monolayer due to their similarities in cell structure. Therefore, the use of Caco-2 cells in a co-culture with RAW264.7 macrophages represents a promising alternative to animal models for predicting the bioactive compounds associated with intestinal IBDs [7,8].
Dietary ingredients, including flavonoids and phenolic chemicals, prevent TJ damage [2,9]. It has been implied that treatment with dietary bioactive compounds has the potential to prevent IBD [10]. Therefore, natural anti-inflammatory compounds, compared to conventional nonsteroidal anti-inflammatory drugs, are considered valuable based on their effective suppression of inflammation without causing side effects or inducing gastrointestinal toxicity [11].
Natural products, including those derived from Dendrobium, are recognized as sources of plant-based alkaloids and are in increasingly high demand by consumers and the food industry [12]. Dendrobium is a genus in the Orchidaceae family and is currently widely cultivated in Asia as a functional food source [13]. Dendrobine is an active alkaloid compound that has been found in Dendrobium [14]. Previous studies have shown that dendrobine can regulate biological functions due to its antitumor, antioxidant, and anti-inflammatory activities [15]. However, there are few reports of the effects of dendrobine on intestinal health, and thus an explanation of how dendrobine exerts its anti-inflammatory effect in this context is lacking. Therefore, we focused on the use of alkaloids to distinguish prevention from inflammation.
Here, we established a co-culture model, as follows. RAW264.7 macrophages were stimulated with Lipopolysaccharides (LPS) to induce inflammation, and Caco-2 cells were exposed to dendrobine. To investigate the regulation of the epithelial barrier by dendrobine, we examined the ability of dendrobine to increase Transepithelial Electrical Resistance (TEER) in the apical Caco-2 cell monolayer and evaluated changes in TJ protein levels by western blot. Furthermore, levels of inflammatory markers and Nuclear Factor Kappa B (NF-κB) pathway activation, which are both related to IBD, were evaluated following macrophage activation to assess the anti-inflammatory effects of dendrobine.
2. MATERIALS AND METHODS
2.1. Materials
A dendrobine standard (purity, >98%) was obtained from the Shanghai Yuanye Biology Technique Co. Ltd. (Shanghai, China), and analytical-grade solvents and LPS were obtained from Sigma-Aldrich (Oakville, ON, Canada). Dulbecco’s Modified Eagle Medium (DMEM) was obtained from Sigma-Aldrich, and Fetal Bovine Serum (FBS) and gentamycin were obtained from Gibco (Grand Island, NE, USA). Enzyme-linked Immunosorbent Assay (ELISA) kits for Nitric Oxide (NO), Tumor Necrosis Factor (TNF)-α, Interleukin (IL)-1β, IL-6, and Prostaglandin E (PGE2) were purchased from the Nanjing Jiancheng Institute of Biotechnology (Nanjing, China). RNAiso Plus (TRIzol) was obtained from Takara (Cat. #9109; Shiga, Japan). Western blot analysis was performed using the following antibodies: anti-phosphorylated (p)-p65 (Cat. #3033), anti-p65 (Cat. #8242), anti-p-IκBα (Cat. #2859), and anti-IκBα (Cat. #4812) from Cell Signaling Technology (Danvers, MA, USA). Anti-occludin (Cat. #13409-1-ap) and anti-ZO-1 (Cat. #21773-1-ap) were obtained from Proteintech (Rosemont, IL, USA). A BCA kit (AR0146) was obtained from Boster Biological Technology (Pleasanton, CA, USA). Polyvinylidene Difluoride (PVDF) membranes were obtained from Millipore (Merck KGaA, Darmstadt, Germany).
2.2. Cell Culture
The experiment was conducted according to the methods of a previous report [10], with some modifications. Caco-2 and RAW264.7 cells were obtained from the Shanghai Cell Bank/Stem Cell Bank at the Chinese Academy of Sciences (Shanghai, China). The cell line culture condition had a 5% CO2 atmosphere using DMEM, with 10% (v/v) FBS (Gibco). The RAW264.7 macrophages were seeded at a density of 5 × 105 cells per cm2, while the Caco-2 cells were plated at density of 5 × 105 cells per cm2 on polycarbonate membranes (MillicellTM Culture Plate Inserts, 12 mm diameter) with a pore size of 0.4 μm (Millipore). All experiments were performed on differentiated Caco-2 cell monolayers at 21 days and a 24 h culture of RAW264.7 macrophages post-seeding.
2.3. Sodium Fluorescein Assay
A sodium fluorescein assay was used to verify the tightness of the Caco-2 monolayers. It was performed according to the methods of a previous study [16].
2.4. TEER Measurement
Transepithelial electrical resistance was measured by using a Millipore ERS-2 resistor (Millipore). Only monolayers with TEER values >800 Ω·cm2 were used in the experiments [7]. TEER values were measured following different treatments according to a previously described method [13].
2.5. Cell Viability Assay
Cell viability was measured at 24 h using an MTT assay to evaluate dendrobine cytotoxicity to Caco-2 and RAW264.7 cells according to a previously described method [13].
2.6. Intestinal Anti-inflammatory Experiments
To determine the intestinal anti-inflammatory potential of dendrobine, the Caco-2 cell monolayers were pre-seeded for 21 days on the apical side and the RAW264.7 macrophages were pre-seeded for 24 h on the basolateral side in transwell plates. The experiment proceeded with the addition of the dendrobine standard (1 and 10 μg/mL) to the apical side of the co-culture system at 37°C for 2 h. The RAW264.7 cells on the basolateral side were then stimulated by the addition of 1 μg/mL LPS for 3 h, according to a previously described method [10]. At the end of the incubation period, the basolateral culture media were collected for subsequent measurement. Caco-2 and RAW264.7 cells were then used for subsequent analysis.
2.7. Measurement of Cytokines, PGE2, and NO Secretion
Levels of IL-6, IL-1β, TNF-α, and PGE2 in the basolateral culture media were quantified using the ELISA kits according to the manufacturer’s instructions. The NO level of the basolateral culture media was also quantified using the Griess method. Quantification was performed after calibration to the standards.
2.8. Quantitative Real-Time Polymerase Chain Reaction
Total RNA was isolated from cells using TRIzol reagent according to the manufacturer’s instructions. Equal amounts of total RNA from each sample were reverse transcribed to cDNA. The sequences of the primers are provided in Table 1. The reaction mixtures were incubated for an initial denaturation at 50°C for 3 min, followed by 95°C for 10 min and 40 cycles of 10 s at 95°C and 30 s at 60°C. The relative amount of each gene was calculated using the 2−ΔΔCT method.
| Primer | Primer sequences (5′→3′) | NCBI reference |
|---|---|---|
| iNOS-mF | GTTCTCAGCCCAACAATACAAGA | NM 010927.4 |
| iNOS-mR | GTGGACGGGTCGATGTCAC | |
| COX2-mF | TTCCAATCCATGTCAAAACCGT | NM 011198.4 |
| COX2-mR | AGTCCGGGTACAGTCACACTT | |
| IL-1β-mF | GAAATGCCACCTTTTGACAGTG | NM 008361.4 |
| IL-1β-mR | TGGATGCTCTCATCAGGACAG | |
| IL-6-mF | TAGTCCTTCCTACCCCAATTTCC | NM 031168.2 |
| IL-6-mR | TTGGTCCTTAGCCACTCCTTC | |
| TNF-α-mF | CTGAACTTCGGGGTGATCGG | XM 021149735.1 |
| TNF-α-mR | GGCTTGTCACTCGAATTTTGAGA | |
| GAPDH-mF | GGTGAAGGTCGGTGTGAACG | XM 017321385.2 |
| GAPDH-mR | CTCGCTCCTGGAAGATGGTG |
GAPDH, Glyceraldehyde-3-phosphate dehydrogenase.
Primer sequences
2.9. Western Blot
Total protein was extracted using radioimmunoprecipitation assay buffer, and the protein concentrations were determined using the BCA kit. Proteins (20 μg) were separated and subsequently transferred to PVDF membranes. The membranes were blocked with 5% bovine serum albumin containing 0.05% Tween-20 in Tris-buffered saline for 4 h at room temperature, followed by incubation with primary antibodies overnight at 4°C. The membranes were then washed twice and incubated with secondary antibodies for 1 h at room temperature. β-actin was used as the loading control. Protein bands were detected by enhanced chemiluminescence.
2.10. Statistical Analysis
All values are expressed as the mean ± Standard Deviation (SD). Statistical analyses were performed using GraphPad Prism (version 7.03; San Diego, CA, USA). The results were analyzed via one-way Analysis of Variance (ANOVA) multiple comparisons; *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, and NS means non-significant as compared with the control.
3. RESULTS
3.1. Establishment of Caco-2 Monolayers
The intestinal-like model was first established and verified before conducting subsequent experiments. TEER values increased with time following incubation, with all values at ~800 Ω·cm2 after the 21-day incubation, which suggested the formation of a TJ structure (Supplementary Figure S1A). Verification of Caco-2 monolayer formation indicated a sodium fluorescein absorptivity within 0–5% (Supplementary Figure S1B), confirming its formation according to international standard terminology [13]. Furthermore, we observed no dendrobine cytotoxicity at 1 or 10 µg/mL (Supplementary Figure S1C). Hence, according to the tests above, the Caco-2 monolayers were formed successfully, and were subsequently prepared for establishing the co-culture model.
3.2. Dendrobine Supports Intestinal Integrity
A previous study indicated that pathogenesis of IBD increased intestinal permeability [4]. In the co-culture system, the LPS treatment group showed a 0.81-fold increase in TEER relative to untreated controls (p < 0.05). The result showed that the intestinal inflammatory model was formed, leading to high intestinal permeability. Additionally, dendrobine treatment protected the Caco-2 cell monolayer, as indicated by the fact that TEER values in the dendrobine treatment group were similar to those in controls (Figure 1A). The TEER values reflected that intestinal permeability was low after exposure with dendrobine.
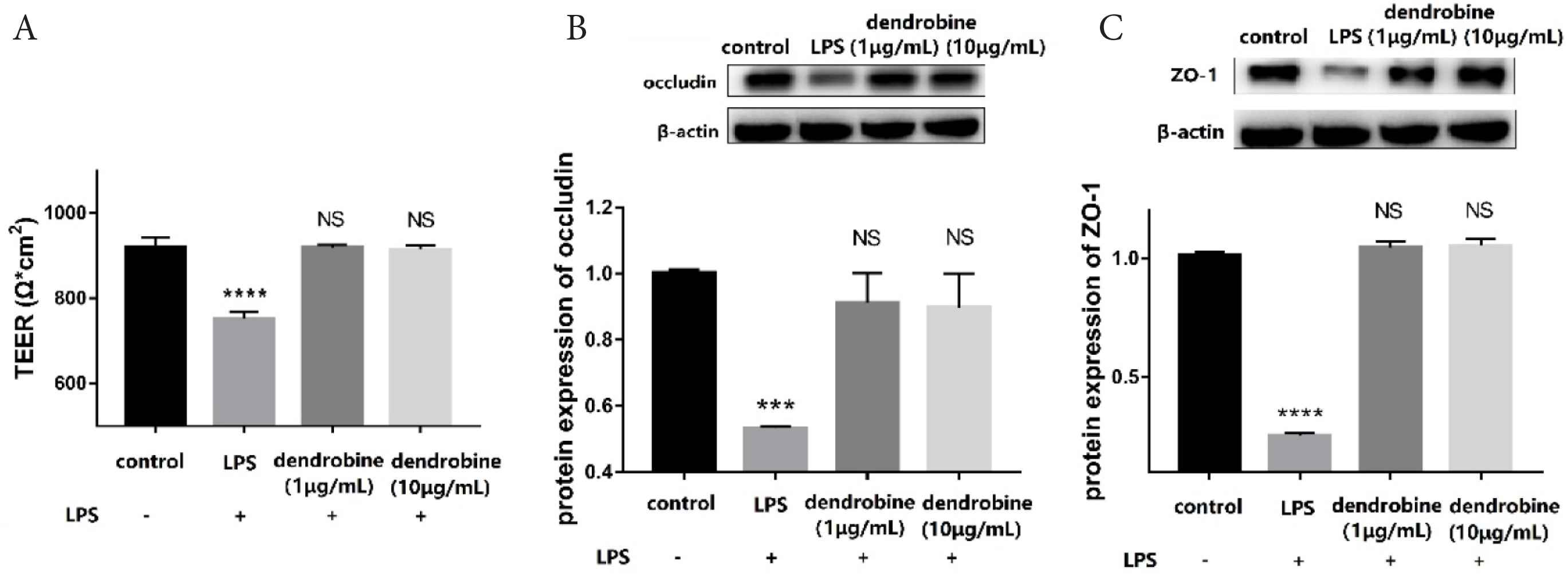
Dendrobine effects on TJs. (A) TEER values and (B and C) western blot results (n = 4). Levels of (B) occludin, (C) ZO-1 in Caco-2 cells (apical side) in the co-culture model. Each value represents the mean ± SD. The results were analyzed via one-way ANOVA multiple comparisons; ***p < 0.001, ****p < 0.0001, NS: non-significant was compared with control.
The proteins forming the tight junctions between adjacent IECs comprise the basic unit of the intestinal epithelial barrier [2]. To determine whether dendrobine protects the in vitro Caco-2/RAW264.7 model from LPS-induced disruption through regulation of TJs, we analyzed levels of ZO-1 and occludin in each experimental group. LPS treatment significantly downregulated occludin levels (p < 0.05), whereas dendrobine treatment inhibited this effect in Caco-2 cells (Figure 1B). Moreover, LPS treatment significantly decreased ZO-1 levels (p < 0.05) relative to the control, whereas dendrobine treatment significantly increased ZO-1 levels in Caco-2 cells (p < 0.05; Figure 1C). Dendrobine treatment at 1 and 10 μg/mL showed no significant difference with the control in ZO-1 and occludin expression (p > 0.05). These results are in agreement with those of previous reports that showed that bioactive compounds protect intestinal integrity by up-regulating the TJ proteins [9,17]. Therefore, a protective effect of dendrobine can be concluded, based on its regulation of TJ protein levels in Caco-2 cells.
3.3. Dendrobine Inhibits NO and PGE2 Production
In inflammation, macrophages are activated, leading a series of mediator levels, such as NO and PEG2, to increase abnormally [18]. To investigate the potential anti-inflammatory effects of dendrobine, NO and PGE2 production and their relative protein expression was determined. Inducible Nitric Oxide Synthase (iNOS) catalyzes NO production [19], which suggests that inhibition of excess NO production and expression of iNOS could be a therapy to prevent IBD. The results indicated that dendrobine (10 μg/mL) inhibited NO production (Figure 2A) based on a 0.84-fold decrease in NO level relative to the LPS group. Additionally, when treating with dendrobine (1 and 10 μg/mL) on the apical side, NO levels on the basolateral side were comparable with control levels (Figure 2A). As seen in Figure 2B, the levels of iNOS mRNA expression levels changed in a negative dose-dependent manner to the dendrobine concentration. Moreover, we found that dendrobine (10 μg/mL) resulted in a decrease in iNOS expression relative to that observed in LPS-treated cells (p < 0.05; Figure 2B). A likely explanation for the observed decrease in NO level could be concomitant downregulation of iNOS expression by LPS-activated macrophages.
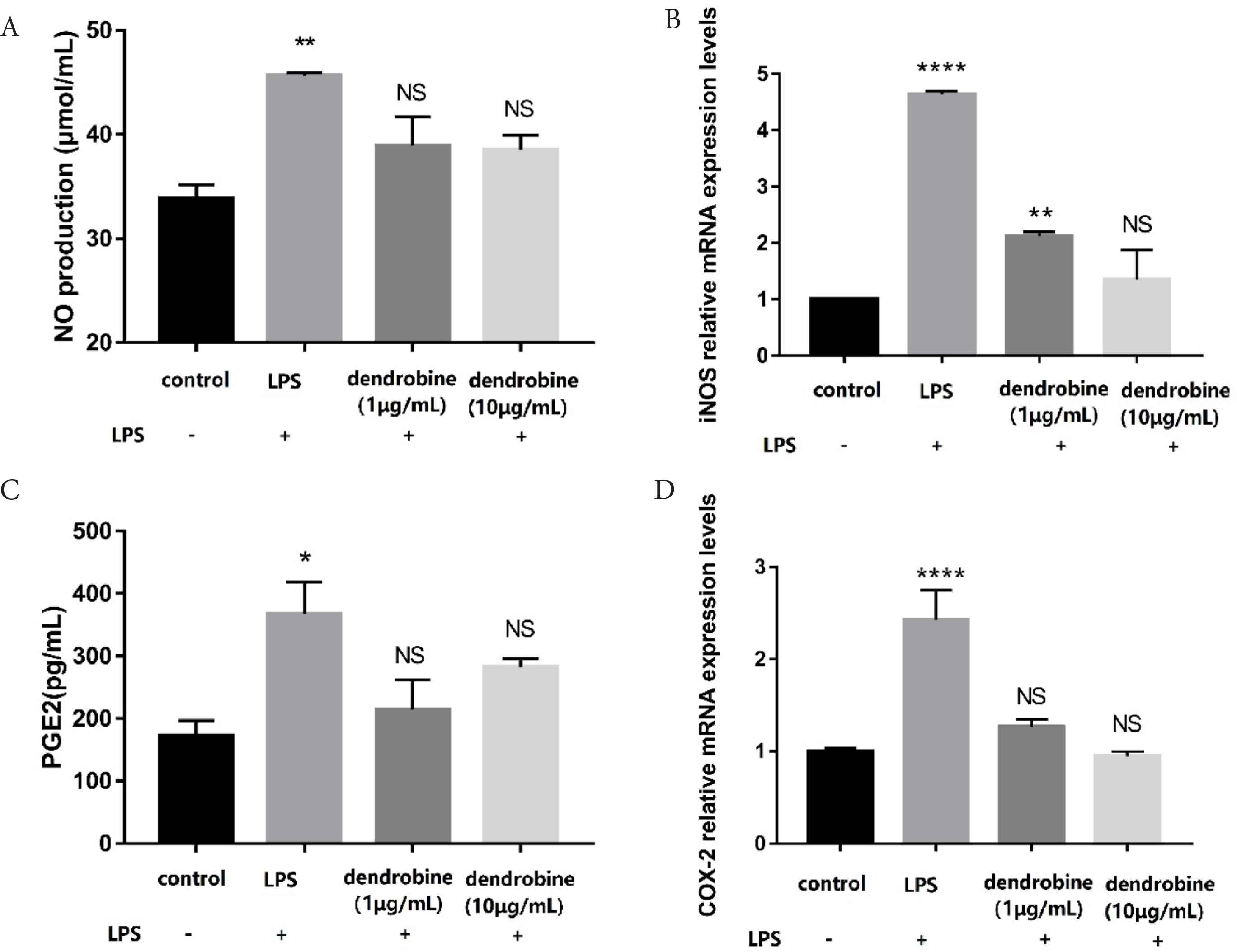
The effect of dendrobine on NO and PGE2 production and iNOS and COX-2 mRNA expression. (A) NO and (C) PGE2 production in co-cultured RAW264.7 macrophages (basolateral side) according to ELISA (n = 4). (B) iNOS and (D) COX-2 expression in co-cultured RAW264.7 macrophages (basolateral side) according to RT-PCR (n = 4). Each value represents the mean ± SD. The results were analyzed via one-way ANOVA multiple comparisons; *p < 0.05, **p < 0.01, ****p < 0.0001, NS: non-significant was compared with control.
Cyclooxygenase-2 (COX-2), induced by inflammatory stimulation, catalyzes arachidonic acid to produce prostaglandin, resulting in the secretion of large amounts of PGE2 and leading to intestinal inflammation [10]. On the basolateral side, the dendrobine group resulted in a decrease in PGE2 level relative to that observed in LPS-treated cells (p < 0.05; Figure 2C). We found that dendrobine treatment exhibited anti-inflammatory activity according to attenuated COX-2 expression (Figure 2D). A previous study indicated that decreasing NO and PGE2 levels could be a way to alleviate inflammation [20]. The decrease in NO and PGE2 after treatment with dendrobine reflected an anti-inflammatory effect.
3.4. Dendrobine Inhibits Cytokine Production
Excess production of proinflammatory cytokines in macrophages may increase the permeability of the intestinal epithelium [1]. After LPS stimulation of RAW264.7 macrophages to mimic intestinal inflammation, we measured levels of proinflammatory cytokines (IL-1β, IL-6, and TNF-α) following dendrobine treatment (Figure 3). We found significantly decreased levels of TNF-α content, and the mRNA levels were lower in LPS-stimulated RAW264.7 cells treated with dendrobine (p < 0.05; Figure 3A and 3C). The 0.52-fold decrease in IL-1β mRNA expression in the dendrobine group (10 μg/mL) compared to the LPS group is shown in Figure 3F. Additionally, we observed that dendrobine (10 μg/mL) treatment dramatically decreased the IL-6 level compared to the LPS group in activated RAW264.7 cells (Figure 3B and 3E). There are some reports on the intestinal anti-inflammatory activity of alkaloids such as berberine [21] or oxymatrine [22] in decreasing the level of cytokines. Similarly, the results of the current study demonstrated that the addition of dendrobine in the apical side attenuated proinflammatory cytokine production in the basolateral side of the co-culture system.
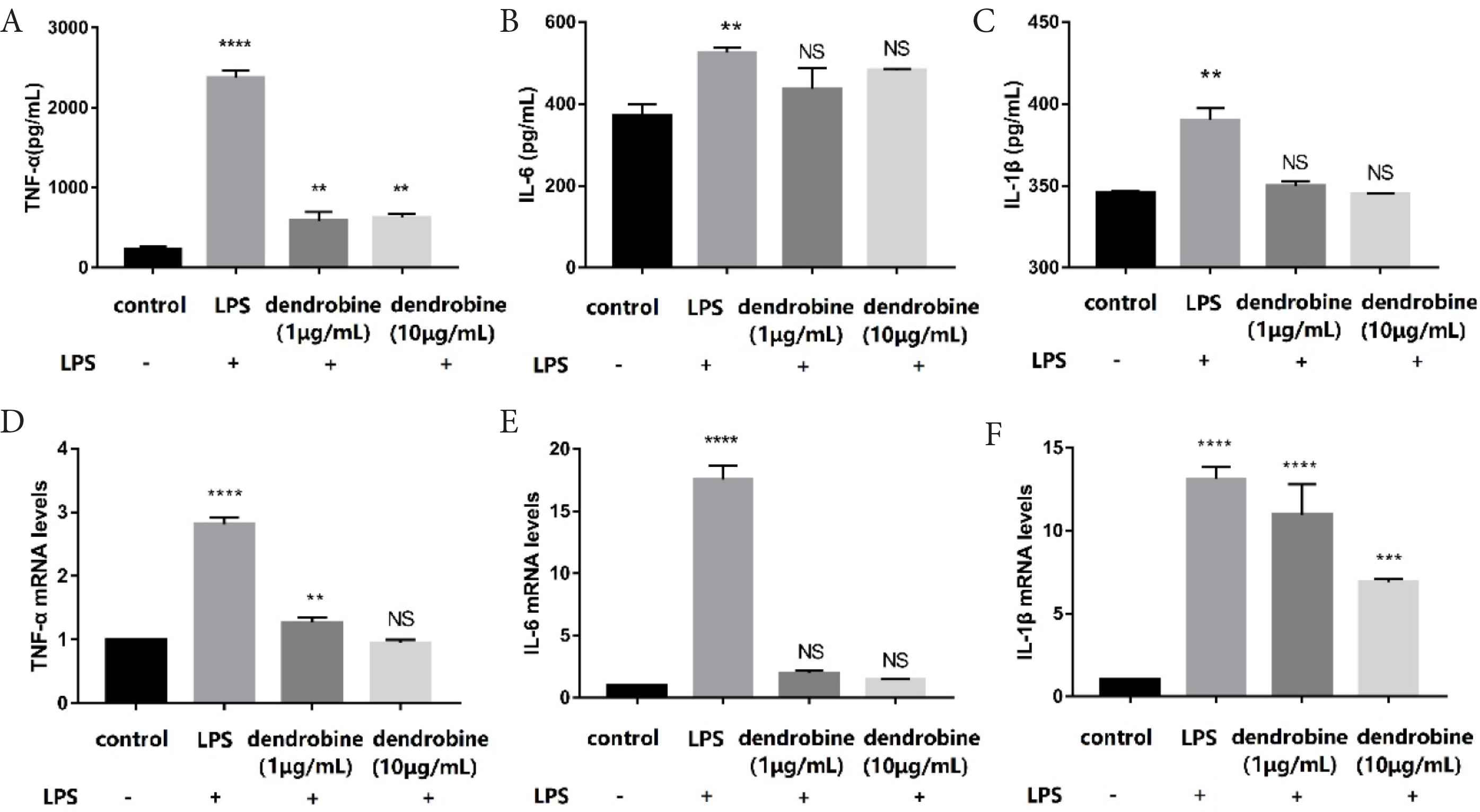
The effect of dendrobine on cytokine production (basolateral side). (A–C) Content and (D–F) mRNA levels of cytokines in RAW264.7 macrophages in the co-culture according to ELISA and qRT-PCR analyses, respectively (n = 4). Each value represents the mean ± SD. The results were analyzed via one-way ANOVA multiple comparisons; **p < 0.01, ***p < 0.001, ****p < 0.0001, NS: non-significant was compared with control.
3.5. Dendrobine Inhibits the NF-κB Signaling Pathway in RAW264.7 Cells of the Co-culture Model
Nuclear factor kappa B regulates proinflammatory mediators in stimulated macrophages causing subsequent upregulation of iNOS, COX-2, and proinflammatory cytokines such as TNF-α. Additionally, activation of NF-κB is associated with the hyperphosphorylation and degradation of IκBα [20]. To elucidate the molecular mechanism of the dendrobine-mediated regulation of inflammation at the intestinal epithelial barrier, we evaluated its effect on the NF-κB pathway. We observed upregulated phosphorylation of the p65 subunit and IκB (according to increased p-p65/p65 and p-IκB/IκB ratios) in LPS-stimulated RAW264.7 cells. However, treatment with dendrobine markedly (p < 0.05) decreased these ratios and downregulated the levels of both p-p65 and p-IκB (Figure 4). It is known that ameliorating the protein expression ratio of p-p65/p65 and p-IκB/IκB can suppress the NF-κB pathway [23]. Similarly, boldine is an alkaloid that suppresses IBD by targeting the NF-κB pathway [24]. The results of the current study indicate that dendrobine in the apical side alters the phosphorylation of proteins regulating NF-κB activity of macrophages in the basolateral side, thereby attenuating NF-κB signaling, which could possibly alleviate IBD symptoms.
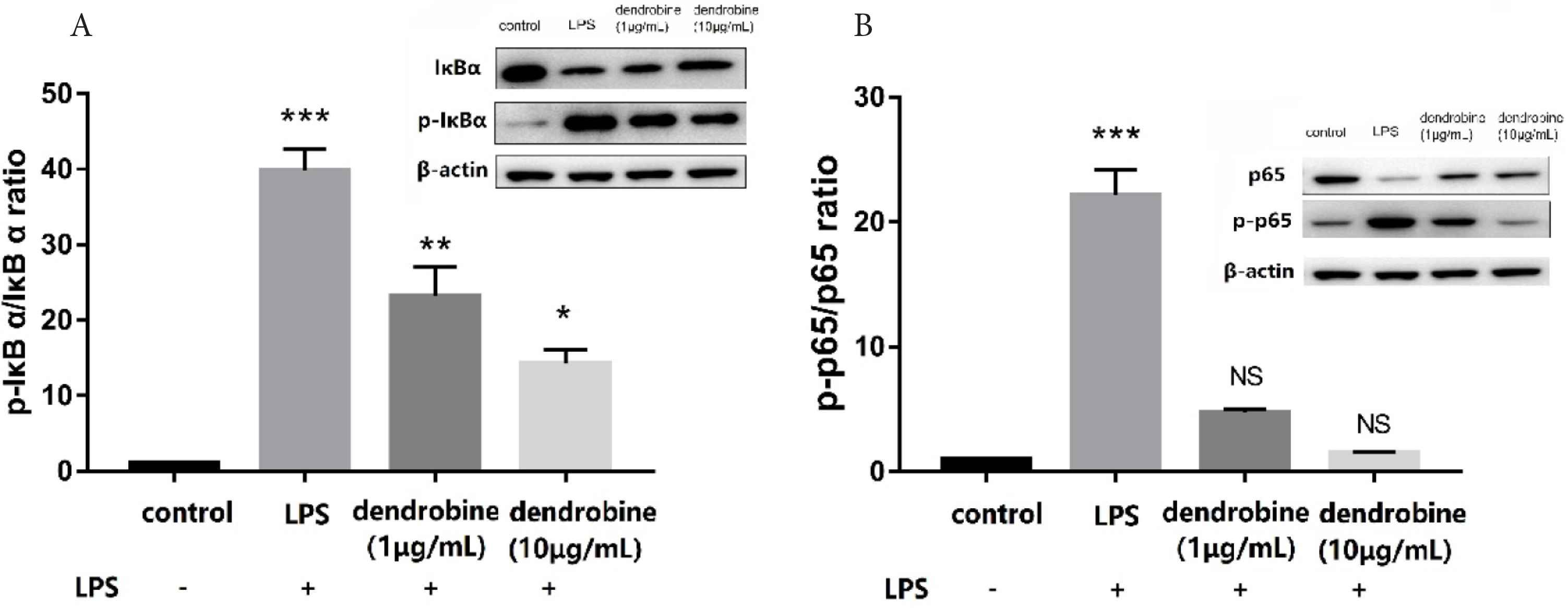
The effect of dendrobine on the NF-κB pathway. Changes in the (A) p-IκBα/IκBα and (B) p-p65/p65 ratios in LPS-stimulated RAW264.7 cells (basolateral side) after dendrobine treatment according to western blot (n = 4). Each value represents the mean ± SD. The results were analyzed via one-way ANOVA multiple comparisons; *p < 0.05, **p < 0.01, ***p < 0.001, NS: non-significant was compared with control.
4. DISCUSSION
As a traditional food and medicine, administration of Dendrobium allows dendrobine to exert its bio-functional effect in the body, especially exposure in gastrointestinal tract. The mechanism through which dendrobine exposure affects the inflamed intestine has hitherto been unclear. Therefore, we elucidated the underlying intestinal anti-inflammatory effect of dendrobine in this study.
Lipopolysaccharides-mediated intestinal inflammatory responses and the resulting damage to the intestinal TJ barrier from LPS are widely reported in studies of IBD. The disruption of TJ protein function induces IBD [17]. TJ proteins, acting as a natural intestinal barrier, prevent toxins and microbial antigens from damaging the lamina propria. A previous study showed that decreases in the levels of TJ proteins contributes to colitis [4]. To determine the TJ of IECs, the TEER was widely applied in the Caco-2 monolayer [25]. In the present study, this co-culture model showed that TEER decreased significantly following basolateral LPS treatment, but that dendrobine treatment upregulated levels of TJ proteins. These results suggest that dendrobine supports intestinal barrier integrity by regulating TJ protein production.
Previous studies have demonstrated that changes in intestinal epithelial monolayer permeability are directly related to the alteration of TJs, and demonstrating the importance of cytokines and TJ proteins in securing the intestinal barrier [2,9]. TNF-α is produced mainly by activated macrophages, monocytes, and T cells and is one of the major cytokines associated with IBD [26]. The imbalance of cytokines may affect inflammation, including IBD [27]. Inhibition of NF-κB activation is an effective strategy to prevent the production of excess cytokines [28]. In this study, the levels of cytokines (IL-6, TNF-α, and IL-1β) and anti-inflammatory mediators (NO and PGE2) decreased in the dendrobine treatment group. Further, we found that LPS treatment increased the phosphorylation of regulators of NF-κB-mediated proinflammatory signaling, whereas dendrobine treatment decreased these levels and inhibited NF-κB activation in RAW264.7 cells while also upregulating TJ protein levels in Caco-2 cells (Figure 5).
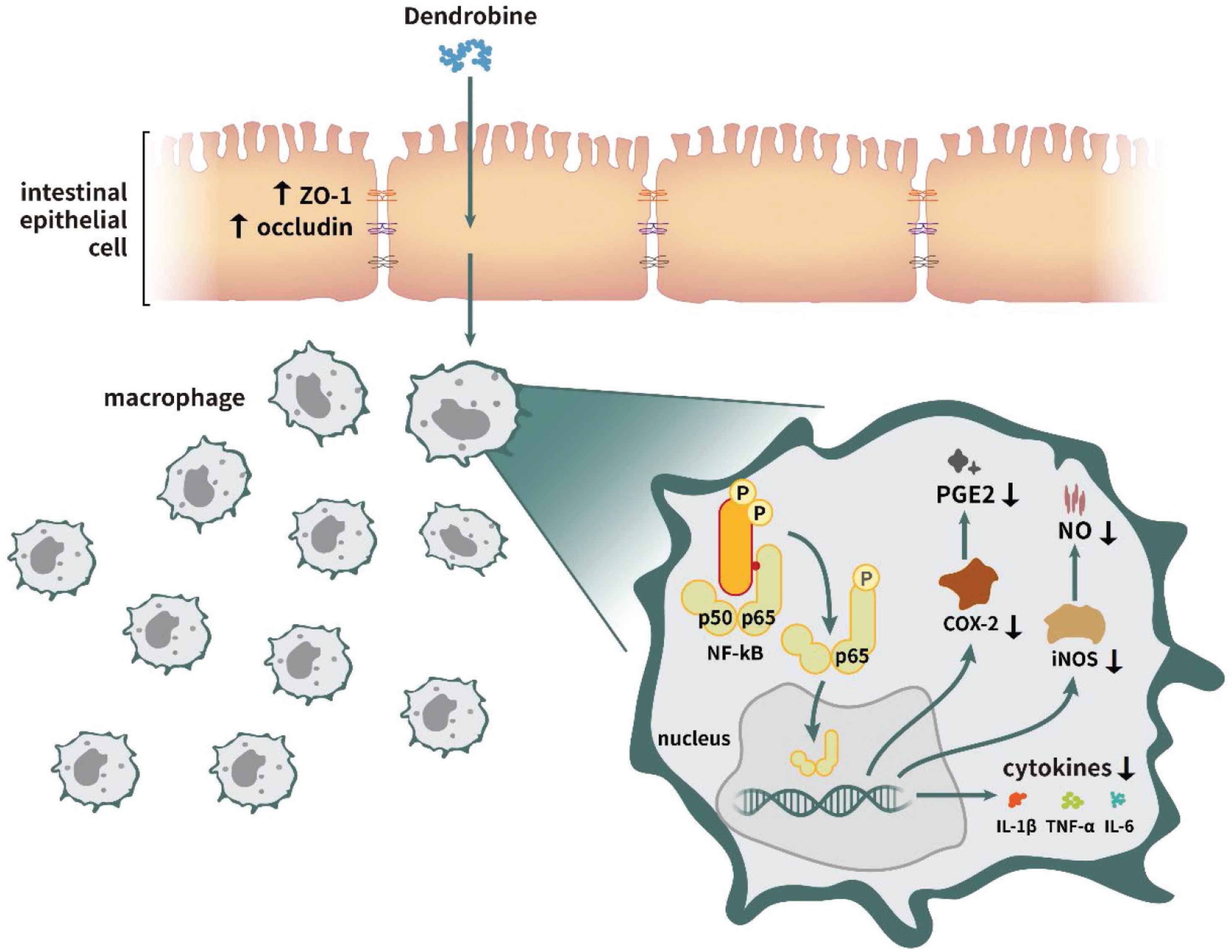
Proposed dendrobine-mediated intestinal anti-inflammatory mechanism.
Similarly, it has been reported that other natural compounds, such as purple carrot anthocyanins, Sambucus nigra fruit extract, and Kaempferol can directly or indirectly mitigate inflammation in RAW264.7 cells through the Caco-2 monolayer [2,7,10]. These examples support our results that dendrobine, as a natural compound, decreases the inflammatory level in the co-culture model. Therefore, dendrobine might directly or indirectly prevent the secretion of proinflammatory cytokines in RAW264.7 macrophages by the NF-κB pathway, and inhibiting protection of TJs between intestinal epithelial cells.
5. CONCLUSION
In summary, these findings indicate that dendrobine, as a natural bioactive compound, may be a potential supplement to prevent IBD-related diseases. Further study will investigate its bio-availability to determine the most effective concentration of dendrobine in vivo. These results will help support the common functional beverage industry and the development of foods containing these bioactive compounds.
CONFLICTS OF INTEREST
The authors declare they have no conflicts of interest.
AUTHORS’ CONTRIBUTION
HL designed the study, analyzed and interpreted the data. JL prepared the paper. LM performed the experiments and interpreted the data. QW provided analysis for the study. GX interpreted the findings.
FUNDING
This study was funded by the
ABBREVIATIONS
- COX-2,
cyclooxygenase-2;
- DMEM,
Dulbecco’s modified Eagle medium;
- ELISA,
enzyme-linked immunosorbent assay;
- FBS,
fetal bovine serum;
- GC,
gas chromatography;
- IBD,
inflammatory bowel disease;
- IEC,
intestinal epithelial cell;
- IL,
interleukin;
- iNOS,
inducible nitric oxide synthase;
- LPS,
lipopolysaccharide;
- MS,
mass spectrometry;
- NF-κB,
nuclear factor kappa B;
- NO,
nitric oxide;
- PVDF,
polyvinylidene difluoride;
- RT-PCR,
polymerase chain reaction;
- TJ,
tight junction;
- TNF,
tumor necrosis factor;
- PGE,
prostaglandin E;
- TEER,
transepithelial electrical resistance;
- ZO-1,
zonula occludens protein 1.
SUPPLEMENTARY MATERIALS
Supplementary data related to this article can be found at
Footnotes
REFERENCES
Cite this article
TY - JOUR AU - Huifan Liu AU - Jiaxi Liang AU - Gengsheng Xiao AU - Lukai Ma AU - Qin Wang PY - 2021 DA - 2021/04/15 TI - Dendrobine Suppresses Lipopolysaccharide-induced Gut Inflammation in a Co-culture of Intestinal Epithelial Caco-2 Cells and RAW264.7 Macrophages JO - eFood SP - 92 EP - 99 VL - 2 IS - 2 SN - 2666-3066 UR - https://doi.org/10.2991/efood.k.210409.001 DO - 10.2991/efood.k.210409.001 ID - Liu2021 ER -